Сколько кислорода способен присоединить один грамм гемоглобина
Обычно из легких в ткани переносятся эритроцитами в химической связи с гемоглобином около 97% кислорода. Оставшиеся 3% кислорода транспортируются в физической растворенной форме плазмой крови. Таким образом, в нормальных условиях почти весь кислород переносится в ткани, будучи связанным с гемоглобином.
Химический состав гемоглобина представлен в наших остальных статьях, где говорилось, что молекула кислорода легко и обратимо связывается с гемом гемоглобина. При высоком Ро2, как это бывает в легочных капиллярах, кислород связывается с гемоглобином, а при низком Р02, как в капиллярах тканей, кислород освобождается от связи с гемоглобином. Такой механизм обеспечивает почти весь транспорт кислорода из легких в ткани.
Кривая диссоциации оксигемоглобина. На рисунке приведена кривая диссоциации оксигемоглобина, демонстрирующая прогрессивный прирост процентной доли оксигемоглобина (процента насыщения гемоглобина кислородом) при увеличении Ро2 в крови. В крови, покидающей легкие и входящей в системные артерии, напряжение О2 обычно составляет примерно 95 мм рт. ст., и на кривой диссоциации видно, что насыщение системной артериальной крови кислородом составляет 97%.
В нормальной возвращающейся из периферических тканей венозной крови напряжение О2 составляет около 40 мм рт. ст. и 75% — насыщение гемоглобина кислородом.
Максимальное количество кислорода, которое может находиться в связи с гемоглобином крови. В 100 мл крови здорового человека содержится около 15 г гемоглобина, и каждый грамм гемоглобина может связать максимально 1,34 мл кислорода (химически чистый гемоглобин может связать 1,39 мл кислорода, но примеси типа метгемоглобина снижают это количество). Итак, 15×1,34 = 20,1, значит, в среднем содержащееся в 100 мл количество гемоглобина при 100% насыщении может связать около 20 мл кислорода. Обычно это обозначают как 20 об% (объемных процентов).
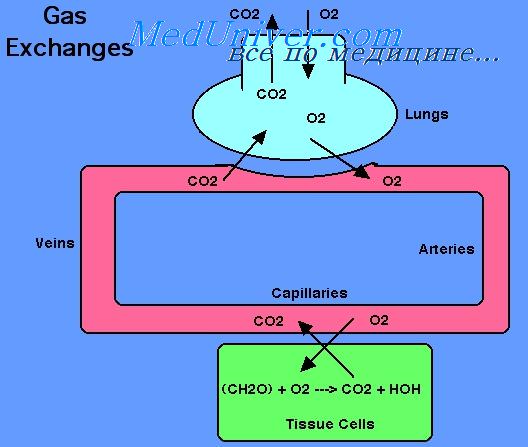
Кривая диссоциации оксигемоглобина может строиться не от процентного насыщения гемоглобина кислородом, а от количества содержания объемных процентов кислорода.
Количество кислорода, высвобождаемого гемоглобином во время прохождения артериальной крови через ткани.
В обычных условиях в системной артериальной крови, насыщенной кислородом на 97%, общее количество связанного с гемоглобином кислорода составляет около 19,4 мл на 100 мл крови. При прохождении через капилляры ткани этот объем снижается до 14,4 мл (Р02 — 40 мм рт. ст., насыщение гемоглобина — 75%). Итак, в нормальных условиях каждые 100 мл крови доставляют от легких к тканям около 5 мл кислорода.
При тяжелой физической работе мышечные клетки потребляют кислород с повышенной скоростью, что может привести к падению Р02 в интерстициальной жидкости мышцы от нормального уровня 40 мм рт. ст. до 15 мм рт. ст.
При таком низком парциальном давлении в каждых 100 мл крови остается только 4,4 мл связанного с гемоглобином кислорода. В этом случае каждые 100 мл протекающей крови отдают тканям 19,4 — 4,4, или 15 мл кислорода, т.е. каждый объем крови отдает тканям в 3 раза больше кислорода, чем в покое. Вспомните, что у хорошо тренированных бегунов-марафонцев сердечный выброс может увеличиться в 6-7 раз, чем при покое.
Если умножить это увеличение сердечного выброса (6-7 раз) на увеличение высвобождения кислорода в тканях каждым объемом крови (3 раза), получается, что к тканям было доставлено в 20 раз больше кислорода, чем в покое. Далее в этой главе Вы узнаете о существовании других факторов, улучшающих доставку кислорода в ткани во время физической нагрузки, поэтому даже при очень напряженной физической работе наблюдается только очень небольшое снижение Ро2 в мышечной ткани.
источник
Транспорт кислорода кровью
Большая часть кислорода в организме млекопитающих переносится кровью в виде химического соединения с гемоглобином. Свободно растворенного кислорода в крови всего 0.3%. Реакцию оксигенации, превращение дезоксигемоглобина в оксигемоглобин, протекающую в эритроцитах капилляров легких можно записать следующим образом:
Эта реакция протекает очень быстро – время полунасыщения гемоглобина кислородом около 3 миллисекунд. Гемоглобин обладает двумя удивительными свойствами, которые позволяют ему быть идеальным переносчиком кислорода. Первое – это способность присоединять кислород, а второе – отдавать его. Оказывается способность гемоглобина присоединять и отдавать кислород зависит от напряжения кислорода в крови. Попробуем изобразить графически зависимость количества оксигенированного гемоглобина от напряжения кислорода в крови, и тогда нам удастся выяснить: в каких случаях гемоглобин присоединяет кислород, а в каких отдает. Гемоглобин и оксигемоглобин неодинаково поглощают световые лучи, поэтому их концентрацию можно определить спектрометрическими методами.
График, отражающий способность гемоглобина присоединять и отдавать кислород называется «Кривая диссоциации оксигемоглобина». По оси абсцисс на этом графике отложено количество оксигемоглобина в процентах ко всему гемоглобину крови, по оси ординат – напряжение кислорода в крови в мм рт. ст.

Рисунок 9А. Кривая диссоциации оксигемоглобина в норме
Рассмотрим график в соответствии с этапами транспорта кислорода: самая высокая точка соответствует тому напряжению кислорода, которое наблюдается в крови легочных капилляров – 100 мм рт.ст. (столько же, сколько и в альвеолярном воздухе). Из графика видно, что при таком напряжении весь гемоглобин переходит в форму оксигемоглобина – насыщается кислородом полностью. Попробуем рассчитать, сколько кислорода связывает гемоглобин. Один моль гемоглобина может связать 4 моля О2 , а 1грамм Нв связывает 1,39 мл О2 в идеале, а на практике 1,34 мл . При концентрации гемоглобина в крови, например, 140 г/литр количество связанного кислорода составит 140 × 1,34 = 189,6 мл/литр крови. Количество кислорода, которое может связать гемоглобин при условии его полного насыщения, называется кислородной емкостью крови (КЕК). В нашем случае КЕК = 189,6 мл.
Обратим внимание на важную особенность гемоглобина – при снижении напряжения кислорода в крови до 60 мм рт.ст, насыщение практически не изменяется – почти весь гемоглобин присутствует в виде оксигемоглобина. Эта особенность позволяет связывать максимально возможное количество кислорода при снижении его содержания в окружающей среде (например, на высоте до 3000 метров).
Кривая диссоциации имеет s – образный характер, что связано с особенностями взаимодействия кислорода с гемоглобином. Молекула гемоглобина связывает поэтапно 4 молекулы кислорода. Связывание первой молекулы резко увеличивает связывающую способность, так же действуют и вторая, и третья молекулы. Этот эффект называется кооперативное действие кислорода
Артериальная кровь поступает в большой круг кровообращения и доставляется к тканям. Напряжение кислорода в тканях, как видно из таблицы 2, колеблется от 0 до 20 мм рт. ст., незначительное количество физически растворенного кислорода диффундирует в ткани, его напряжение в крови снижается. Снижение напряжения кислорода сопровождается диссоциацией оксигемоглобина и освобождением кислорода. Освободившийся из соединения кислород переходит в физически растворенную форму и может диффундировать в ткани по градиенту напряжения.. На венозном конце капилляра напряжение кислорода равно 40 мм.рт.ст, что соответствует примерно 73% насыщения гемоглобина. Крутая часть кривой диссоциации соответствует напряжению кислорода обычному для тканей организма – 35 мм рт.ст и ниже.
Таким образом, кривая диссоциации гемоглобина отражает способность гемоглобина присоединять кислород, если напряжение кислорода в крови высоко, и отдавать его при снижении напряжения кислорода.
Переход кислорода в ткани осуществляется путем диффузии, и описывается законом Фика, следовательно зависит от градиента напряжений кислорода.
Можно узнать, сколько кислорода извлекается тканью. Для этого нужно определить количество кислорода в артериальной крови и в венозной крови, оттекающей от определенной области. В артериальной крови, как нам удалось вычислить (КЕК) содержится 180-200 мл. кислорода. Венозная кровь в состоянии покоя содержит около 120 мл. кислорода. Попробуем рассчитать коэффициент утилизации кислорода: 180 мл. — 120 мл. = 60 мл.- это количество извлеченного тканями кислорода, 60мл./180 ´ 100 = 33%. Следовательно, коэффициент утилизации кислорода равен 33% (в норме от 25 до 40%). Как видно из этих данных, не весь кислород утилизируется тканями. В норме в течение одной минуты к тканям доставляется около 1000 мл. кислорода. Если учесть коэффициент утилизации, становится ясно, что ткани извлекают от 250 до 400 мл. кислорода в минуту, остальной кислород возвращается к сердцу в составе венозной крови. При тяжелой мышечной работе коэффициент утилизации повышается до 50 – 60 %.
Однако количество кислорода, которое получают ткани, зависит не только от коэффициента утилизации. При изменении условий во внутренней среде и тех тканях, где осуществляется диффузия кислорода, свойства гемоглобина могут измениться. Изменение свойств гемоглобина отражается на графике и называется «сдвиг кривой». Отметим важную точку на кривой – точка полунасыщения гемоглобина кислородом наблюдается при напряжении кислорода 27 мм рт. ст., при таком напряжении 50 % гемоглобина находится в форме оксигемоглобина, 50% в виде дезоксигемоглобина, следовательно 50 % связанного кислорода – свободно (примерно 100мл/л). Если в ткани увеличивается концентрация углекислого газа, ионов водорода, температура, то кривая сдвигается вправо. В этом случае точка полунасыщения переместится к более высоким значениям напряжения кислорода — уже при напряжении 40 мм рт. ст. будет освобождено 50 % кислорода (рисунок 9Б). Интенсивно работающей ткани гемоглобин отдаст кислород легче. Изменение свойств гемоглобина обусловлены следующими причинами: закисление среды в результате увеличения концентрации углекислого газа действует двумя путями 1) увеличение концентрации ионов водорода способствует отдаче кислорода оксигемоглобином потому, что ионы водорода легче связываются с дезоксигемоглобином, 2) прямое связывание углекислого газа с белковой частью молекулы гемоглобина уменьшает ее сродство к кислороду; увеличение концентрации 2,3-дифосфоглицерата, который появляется в процессе анаэробного гликолиза и тоже встраивается в белковую часть молекулы гемоглобина и снижает его сродство к кислороду.
Сдвиг кривой влево наблюдается, например, у плода, когда в крови определяется большое количество фетального гемоглобина.

Рисунок 9 Б. Влияние изменения параметров внутренней среды
Не нашли то, что искали? Воспользуйтесь поиском:
источник
V.1. Гемоглобин и его участие в транспорте кислорода
Основная функция гемоглобина — транспорт кислорода, которую онвыполняет очень эффективно. Гемоглобин артериальной крови насыщен кислородом приблизительно на 96%, гемоглобин венозной крови — на 64% (White А. et al, 1981).
Уменьшение степени насыщения на 32 % соответствует количеству О2, потребляемому тканями. Принимая, что 100 мл крови содержат 15,0 г гемоглобина и что каждый грамм гемоглобина может связать 1,34 мл О2 (число Хюфнера) находим, что:
0,96 х 1,34 х 15 = 19,2 мл О2 —
транспортируется 100 мл артериальной крови (19,2 объемных %);
0,64 х 1,34 х 15 = 12,8 мл О2 —
транспортируется 100 мл венозной крови (12,8 объемных %);
19,2 — 12,8 = 6,4 мл О2 (6,4% объемных %)
0,32 х 1,34 х 15 = 6,4 мл О2 (6,4 % объемных %)
отдается тканям каждыми 100 мл артериальной крови, протекающими через капилляры.
Поскольку минутный объем крови (МОК) равен 5 литрам, в сутках 1440 минут, определяем, что в течение суток общий объем перекачиваемой сердцем крови составит:
А так как один литр артериальной крови способен транспортировать 19,2 объемных % кислорода, из которых 6,4 объемных процента потребляются тканями, находим:
———————— = 1382 литра — количество кислорода, транспортируемое
100 за сутки артериальной кровью
———————— = 461 литр — количество кислорода, потребляемое тканями
100 за сутки из артериальной крови
В целом, за сутки кровь переносит от легких к тканям около 1400 л кислорода, из которых 450-600 л потребляются тканями. Причем, практически весь перенос кислорода обеспечивается гемоглобином, т.к. кислород плохо растворим в водных растворах. Ограниченная растворимость кислорода позволяет транспортировать его в растворенном состоянии в количестве, составляющем всего 0,3 мл О2 / 100 мл крови (White А. еt al, 1981) при РаО2 равном 12,7 кПа (95 мм рт ст). Это соотношение вытекает из закона Генри-Дальтона, в соответствии с которым «количество растворенного в жидкости газа пропорционально его напряжению». Важно отметить, что несмотря на малое количество растворенного кислорода, именно эта его фракция обусловливает РаО2.
Гемоглобин растворен в водном цитозоле эритроцитов в очень высокой концентрации, порядка 34%. Его синтез осуществляется в процессе созревания ретикулоцитов и перехода их в эритроциты. Гемоглобин состоит из белка — глобина и простетической группы — гема. Гем представляет собой соединение, в молекулу которого входят атом двухвалентного железа и 4 замещенных пиррольных кольца, связанных между собой метиновыми группами ( — СН = ). Скелетом молекулы гема служит порфирин. Fe 2+ имеет в геме координационное число, равное 6, т.е. 6 пар электронов. Четыре из этих пар связаны с атомами азота, одна пара с белком (глобином) через гистидин. Оставшаяся пара электронов используется для образования донорско-акцепторной координационной связи (ковалентной химической связи) с кислородом (рис. 12).
Функциональные свойства гемоглобина, обусловленные его уникальной структурой, специфически соответствуют задачам обеспечения дыхательной функции. Каждая из четырех полипептидных цепей глобина содержит по одной молекуле гема. А поскольку атом железа в геме может обратимо связывать одну молекулу О2 понятно, что одна молекула полностью оксигенированного гемоглобина (НbО2) содержит четыре молекулы кислорода.Гемоглобин, отдавший кислород тканям, называется свободным (неоксигенированным) гемоглобином.

Рис. 12.Координационные связи железа в молекуле гема
В процессе присоединения и отдачи кислорода железо в молекуле гемоглобина свою валентность не меняет, т.е. при присоединении кислорода или при его отдаче железо не окисляется и не восстанавливается. Следовательно, в этих случаях речь идет об оксигенированном (оксигемоглобин) и неоксигенированном (свободном) гемоглобине, но не об окисленном и восстановленном гемоглобине.
Окисленный гемоглобин — это метгемоглобин (MetHb), содержащий Fe 3+ . Он не способен присоединять кислород, однако Fe 3+ в MetHb может взаимодействовать со многими анионами, например, при щелочном рН с ОН — и при кислом рН — с Cl — . В небольшом количестве, составляющем 0, 5% от всего гемоглобина, метгемоглобин образуется ежесуточно в условиях in vivo (White A. et al, 1981).
Кроме кислорода к атому железа в молекуле гемоглобина могут присоединяться и другие вещества (лиганды) : Н2О; СО; СN. Их сродство, проявляемое к атому железа, неодинаково. Оно наиболее выражено у цианидов, затем у окиси углерода, затем у кислорода и наименее выражено у воды.
Степень насыщения гемоглобина кислородом зависит от парциального давления кислорода (РО2). Это давление в легких ( 100 мм рт.столба) достаточно, чтобы практически полностью оксигенировать весь гемоглобин в эритроцитах. Уникальной особенностью связывания гемоглобином О2 является кооперативное взаимодействие между гемсвязывающими участками, получившее название гем-гем — взаимодействие. И хотя прямых физических контактов между четырьмя группами гема нет, кооперативность связывания О2 проявляется в том, что по мере протекания оксигенирования гемоглобина облегчается связывание последующих молекул кислорода. При физиологических условиях сродство к О2 у гемоглобина и у полностью оксигенированного гемоглобина отличается в 500 раз (A. White et al, 1981); это показывает, что после присоединения первых молекул О2 сродство частично насыщенного гемоглобина к кислороду увеличивается.Данное положение характеризуетсясигмовидной формой кривой диссоциации HbO2 (рис. 5).Такая формауказывает на то, что связывание О2 одной из гемовых групп гемоглобина влияет на константы диссоциации других гемовых групп этой же молекулы. Сигмовидный характер кривой диссоциации НbО2 имеет большое физиологическое значение. Как видно из рис. 5 парциальное давление О2 существенно влияет на насыщение гемоглобина в пределах от 20 до 60 мм рт. ст., когда показатель насыщения кислородом характеризуется наиболее крутым подъемом отрезка кривой. При больших значениях РО2 кривая насыщения приобретает пологий характер, увеличиваясь в диапазоне 60-90 мм рт. ст. всего на 7%.
Связывание гемоглобином кислорода зависит не только от РО2. Этот процесс тонко регулируется рядом соединений, подобно тому как это имеет место при действии на аллостерический фермент.Таким регуляторным действием обладают 2,3 — дифосфоглицерат, диоксид углерода (СО2) и ионы Н + .
Главной фракцией фосфатов в эритроцитах является 2,3 – дифосфоглицерат (ДФГ); его внутриклеточная концентрация составляет приблизительно 4-5 — ммоль, т.е. сопоставима с концентрацией гемоглобина. Дифосфоглицерат (ДФГ) связывается с тетрамерным гемоглобином при нейтральном рН, образуя комплекс с соотношением компонентов 1 : 1. Связывание с гемоглобином ДФГ обусловлено наличием в молекуле последнего отрицательно заряженных групп (рис. 13):
источник
V.1. Гемоглобин и его участие в транспорте кислорода
Основная функция гемоглобина — транспорт кислорода, которую онвыполняет очень эффективно. Гемоглобин артериальной крови насыщен кислородом приблизительно на 96%, гемоглобин венозной крови — на 64% (White А. et al, 1981).
Уменьшение степени насыщения на 32 % соответствует количеству О2, потребляемому тканями. Принимая, что 100 мл крови содержат 15,0 г гемоглобина и что каждый грамм гемоглобина может связать 1,34 мл О2 (число Хюфнера) находим, что:
0,96 х 1,34 х 15 = 19,2 мл О2 —
транспортируется 100 мл артериальной крови (19,2 объемных %);
0,64 х 1,34 х 15 = 12,8 мл О2 —
транспортируется 100 мл венозной крови (12,8 объемных %);
19,2 — 12,8 = 6,4 мл О2 (6,4% объемных %)
0,32 х 1,34 х 15 = 6,4 мл О2 (6,4 % объемных %)
отдается тканям каждыми 100 мл артериальной крови, протекающими через капилляры.
Поскольку минутный объем крови (МОК) равен 5 литрам, в сутках 1440 минут, определяем, что в течение суток общий объем перекачиваемой сердцем крови составит:
А так как один литр артериальной крови способен транспортировать 19,2 объемных % кислорода, из которых 6,4 объемных процента потребляются тканями, находим:
———————— = 1382 литра — количество кислорода, транспортируемое
100 за сутки артериальной кровью
———————— = 461 литр — количество кислорода, потребляемое тканями
100 за сутки из артериальной крови
В целом, за сутки кровь переносит от легких к тканям около 1400 л кислорода, из которых 450-600 л потребляются тканями. Причем, практически весь перенос кислорода обеспечивается гемоглобином, т.к. кислород плохо растворим в водных растворах. Ограниченная растворимость кислорода позволяет транспортировать его в растворенном состоянии в количестве, составляющем всего 0,3 мл О2 / 100 мл крови (White А. еt al, 1981) при РаО2 равном 12,7 кПа (95 мм рт ст). Это соотношение вытекает из закона Генри-Дальтона, в соответствии с которым «количество растворенного в жидкости газа пропорционально его напряжению». Важно отметить, что несмотря на малое количество растворенного кислорода, именно эта его фракция обусловливает РаО2.
Гемоглобин растворен в водном цитозоле эритроцитов в очень высокой концентрации, порядка 34%. Его синтез осуществляется в процессе созревания ретикулоцитов и перехода их в эритроциты. Гемоглобин состоит из белка — глобина и простетической группы — гема. Гем представляет собой соединение, в молекулу которого входят атом двухвалентного железа и 4 замещенных пиррольных кольца, связанных между собой метиновыми группами ( — СН = ). Скелетом молекулы гема служит порфирин. Fe 2+ имеет в геме координационное число, равное 6, т.е. 6 пар электронов. Четыре из этих пар связаны с атомами азота, одна пара с белком (глобином) через гистидин. Оставшаяся пара электронов используется для образования донорско-акцепторной координационной связи (ковалентной химической связи) с кислородом (рис. 12).
Функциональные свойства гемоглобина, обусловленные его уникальной структурой, специфически соответствуют задачам обеспечения дыхательной функции. Каждая из четырех полипептидных цепей глобина содержит по одной молекуле гема. А поскольку атом железа в геме может обратимо связывать одну молекулу О2 понятно, что одна молекула полностью оксигенированного гемоглобина (НbО2) содержит четыре молекулы кислорода.Гемоглобин, отдавший кислород тканям, называется свободным (неоксигенированным) гемоглобином.
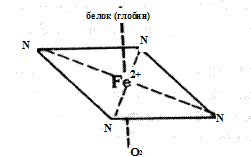
Рис. 12.Координационные связи железа в молекуле гема
В процессе присоединения и отдачи кислорода железо в молекуле гемоглобина свою валентность не меняет, т.е. при присоединении кислорода или при его отдаче железо не окисляется и не восстанавливается. Следовательно, в этих случаях речь идет об оксигенированном (оксигемоглобин) и неоксигенированном (свободном) гемоглобине, но не об окисленном и восстановленном гемоглобине.
Окисленный гемоглобин — это метгемоглобин (MetHb), содержащий Fe 3+ . Он не способен присоединять кислород, однако Fe 3+ в MetHb может взаимодействовать со многими анионами, например, при щелочном рН с ОН — и при кислом рН — с Cl — . В небольшом количестве, составляющем 0, 5% от всего гемоглобина, метгемоглобин образуется ежесуточно в условиях in vivo (White A. et al, 1981).
Кроме кислорода к атому железа в молекуле гемоглобина могут присоединяться и другие вещества (лиганды) : Н2О; СО; СN. Их сродство, проявляемое к атому железа, неодинаково. Оно наиболее выражено у цианидов, затем у окиси углерода, затем у кислорода и наименее выражено у воды.
Степень насыщения гемоглобина кислородом зависит от парциального давления кислорода (РО2). Это давление в легких ( 100 мм рт.столба) достаточно, чтобы практически полностью оксигенировать весь гемоглобин в эритроцитах. Уникальной особенностью связывания гемоглобином О2 является кооперативное взаимодействие между гемсвязывающими участками, получившее название гем-гем — взаимодействие. И хотя прямых физических контактов между четырьмя группами гема нет, кооперативность связывания О2 проявляется в том, что по мере протекания оксигенирования гемоглобина облегчается связывание последующих молекул кислорода. При физиологических условиях сродство к О2 у гемоглобина и у полностью оксигенированного гемоглобина отличается в 500 раз (A. White et al, 1981); это показывает, что после присоединения первых молекул О2 сродство частично насыщенного гемоглобина к кислороду увеличивается.Данное положение характеризуетсясигмовидной формой кривой диссоциации HbO2 (рис. 5).Такая формауказывает на то, что связывание О2 одной из гемовых групп гемоглобина влияет на константы диссоциации других гемовых групп этой же молекулы. Сигмовидный характер кривой диссоциации НbО2 имеет большое физиологическое значение. Как видно из рис. 5 парциальное давление О2 существенно влияет на насыщение гемоглобина в пределах от 20 до 60 мм рт. ст., когда показатель насыщения кислородом характеризуется наиболее крутым подъемом отрезка кривой. При больших значениях РО2 кривая насыщения приобретает пологий характер, увеличиваясь в диапазоне 60-90 мм рт. ст. всего на 7%.
Связывание гемоглобином кислорода зависит не только от РО2. Этот процесс тонко регулируется рядом соединений, подобно тому как это имеет место при действии на аллостерический фермент.Таким регуляторным действием обладают 2,3 — дифосфоглицерат, диоксид углерода (СО2) и ионы Н + .
Главной фракцией фосфатов в эритроцитах является 2,3 – дифосфоглицерат (ДФГ); его внутриклеточная концентрация составляет приблизительно 4-5 — ммоль, т.е. сопоставима с концентрацией гемоглобина. Дифосфоглицерат (ДФГ) связывается с тетрамерным гемоглобином при нейтральном рН, образуя комплекс с соотношением компонентов 1 : 1. Связывание с гемоглобином ДФГ обусловлено наличием в молекуле последнего отрицательно заряженных групп (рис. 13):
источник