Снижают сродство гемоглобина к кислороду в тканях
Молекула гемоглобина может находиться в двух формах — напряженной и расслабленной. Расслабленная форма гемоглобина имеет свойство насыщаться кислородом в 70 раз быстрее, чем напряженная. Изменение фракций напряженной и расслабленной формы в общем количестве гемоглобина в крови обусловливает S-образный вид кривой диссоциации оксигемоглобина, а следовательно, так называемое сродство гемоглобина к кислороду. Если вероятность перехода от напряженной формы гемоглобина к расслабленной больше, то возрастает сродство гемоглобина к кислороду, и наоборот. Вероятность образования указанных фракций гемоглобина изменяется в большую или меньшую сторону под влиянием нескольких факторов.
Основной фактор — это связывание кислорода с геминовой фуппой молекулы гемоглобина. При этом чем больше геминовых фупп гемоглобина связывают кислород в эритроцитах, тем более легким становится переход молекулы гемоглобина к расслабленной форме и тем выше их сродство к кислороду. Поэтому при низком Р02, что имеет место в метаболически активных тканях, сродство гемоглобина к кислороду ниже, а при высоком Р02 — выше. Как только гемоглобин захватывает кислород, повышается его сродство к кислороду и молекула гемоглобина становится насыщенной при связывании с четырьмя молекулами кислорода.

Когда эритроциты, содержащие гемоглобин, достигают тканей, то кислород из эритроцитов диффундирует в клетки. В мышцах он поступает в своеобразного депо кислорода — в молекулы миоглобина, из которого кислород используется в биологическом окислении мышц.
Диффузия кислорода из гемоглобина эритроцитов в ткани обусловлена низким Р02 в тканях — 35 мм рт. ст. Внутри клеток тканей напряжение кислорода, необходимое для поддержания нормального метаболизма, составляет еще меньшую величину — не более 1 кПа. Поэтому кислород путем диффузии из капилляров достигает метаболически активных клеток. Некоторые ткани приспособлены к низкому содержанию Р02 в капиллярах крови, что компенсируется высокой плотностью капилляров на единицу объема тканей. Например, в скелетной и сердечной мышцах Р02 в капиллярах может снизиться чрезвычайно быстро во время сокращения. В мышечных клетках содержится белок миоглобин, который имеет более высокое сродство к кислороду, чем гемоглобин. Миоглобин интенсивно насыщается кислородом и способствует его диффузии из крови в скелетную и сердечную мышцы, где он обусловливает процессы биологического окисления. Эти ткани способны экстрагировать до 70 % кислорода из крови, проходящей через них, что обусловлено снижением сродства гемоглобина к кислороду под влиянием температуры тканей и рН.
Эффект рН и температуры на сродство гемоглобина к кислороду. Молекулы гемоглобина способны реагировать с ионами водорода, в результате этой реакции происходит снижение сродства гемоглобина к кислороду. При насыщении гемоглобина менее 100 % низкое рН понижает связывание кислорода с гемоглобином — кривая диссоциации оксигемоглобина смещается вправо по оси х. Это изменение свойства гемоглобина под влиянием ионов водорода называется эффектом Бора. Метаболически активные ткани продуцируют кислоты, такую как молочная, и С02. Если рН плазмы крови снижается от 7,4 в норме до 7,2, что имеет место при сокращении мыщц, то концентрация кислорода в ней будет возрастать вследствие эффекта Бора. Например, при постоянном рН 7,4 кровь отдавала бы порядка 45 % кислорода, т. е. насыщение гемоглобина кислородом снижалось до 55 %. Однако когда рН снижается до 7,2, кривая диссоциации смещается по оси х вправо. В результате насыщение гемоглобина кислородом падает до 40 %, т. е. кровь может отдавать в тканях до 60 % кислорода, что на 1/з больше, чем при постоянном рН.

Метаболически активные ткани повышают продукцию тепла. Повышение температуры тканей при физической работе изменяет соотношение фракций гемоглобина в эритроцитах и вызывает смещение кривой диссоциации оксигемоглобина вправо вдоль оси х. В результате большее количество кислорода будет освобождаться из гемоглобина эритроцитов и поступать в ткани.
Эффект 2,3-дифосфоглицерата (2,3-ДФГ) на сродство гемоглобина к кислороду. При некоторых физиологических состояниях, например при понижении Р02 в крови ниже нормы (гипоксия) в результате пребывания человека на большой высоте над уровнем моря, снабжение тканей кислородом становится недостаточным. При гипоксии может понижаться сродство гемоглобина к кислороду вследствие увеличения содержания в эритроцитах 2,3-ДФГ. В отличие от эффекта Бора, уменьшение сродства гемоглобина к кислороду под влиянием 2,3-ДФГ не является обратимым в капиллярах легких. Однако при движении крови через капилляры легких эффект 2,3-ДФГ на снижение образования оксигемоглобина в эритроцитах (плоская часть кривой диссоциации оксигемоглобина) выражен в меньшей степени, чем отдача кислорода под влиянием 2,3-ДФГ в тканях (наклонная часть кривой), что обусловливает нормальное кислородное снабжение тканей.
источник
Факторы, влияющие на сродство гемоглобина к кислороду
Динамика кривой зависит от нескольких факторов. Кривая может сдвигаться относительно оси абсцисс вправо или влево (эффект Бора) в зависимости от сопутствующего парциального давления двуокиси углерода и величины рН. При этом реальная физиологическая кривая имеет S-образную форму. При увеличении содержания двуокиси углерода и закис-лении крови кривая диссоциации оксигемоглобина сдвигается вправо и, напротив, при снижении РСо2 и защелачивания крови кривая сдвигается влево.
Биологическое значение кривой диссоциации оксигемоглобина.Кривая диссоциации оксигемоглобина имеет важное биологическое значение для переноса кислорода кровью. Участок кривой, соответствующий низким парциальным значениям кислорода, характеризует содержание оксигемоглобина в капиллярах тканей, а фрагмент кривой, лежащий в области высокого парциального давления кислорода 80—100 мм рт.ст., соответствует крови в легочных капиллярах.
Эффективность транспорта кислородак тканям определяют два фактора: количество оксигемоглобина, образовавшегося в легких, и количество кислорода, отдаваемого тканям, что зависит от степени распада оксигемоглобина в восстановленный гемоглобин. Процесс наиболее эффективного переноса кислорода к тканям соответствует S-образной форме кривой диссоциации оксигемоглобина. В области высокого парциального давления кислорода кривая близка к насыщению кислородом крови, а в области низких значений парциального давления кислорода в тканях значительная часть оксигемоглобина отдает кислород и превращается в восстановленную форму.
Эффект Бора. Огромное биологическое значение для транспорта кислорода и двуокиси углерода имеет эффект Бора.
При увеличении парциального давления двуокиси углерода в тканях кривая диссоциации оксигемоглобина, сдвигаясь вправо, отражает повышение способности оксигемоглобина отдавать кислород тканям и тем самым высвобождаться для дополнительного связывания двуокиси углерода и переноса ее избытка из тканей в легкие.
Напротив, при снижении парциального давления двуокиси углерода и смещении рН крови в основную сторону (алкалоз) сдвиг кривой диссоциации оксигемоглобина влево означает снижение способности оксигемоглобина отдавать кислород тканям и поглощать двуокись углерода для транспорта ее к легким.
Сдвиг кривой диссоциации оксигемоглобина иллюстрирует взаимосвязь транспорта кислорода и двуокиси углерода в крови и сродство гемоглобина к этим газам.
#82 Объясните мех-мы транспорта углекислого газа кровью, роль карбоангидразы. Проанализируйте роль дыхания в регуляции рН крови.
Перенос углекислого газа. Двуокись углерода, образующаяся в тканях, переносится с кровью к легким и выделяется с выдыхаемым воздухом в атмосферу. В отличие от транспорта кислорода она транспортируется кровью тремя способами.
Формы транспорта углекислого газа. Во-первых, так же как и кислород, двуокись углерода переносится в физически растворенном состоянии. Содержание физически растворенной двуокиси углерода в артериальной крови составляет 0,026 мл в 1 мл крови, что в 9 раз превышает количество физически растворенного кислорода. Это объясняется гораздо более высоким коэффициентом растворимости двуокиси углерода.
Во-вторых, двуокись углерода транспортируется в виде химического соединения с гемоглобином — карбогемоглобина.
В третьих — в виде гидрокарбоната НСОз, образующегося в результате диссоциации угольной кислоты.
Механизм переноса двуокиси углерода. Перенос двуокиси углерода из тканей в легкие осуществляется следующим образом. Наибольшее парциальное давление двуокиси углерода в клетках тканей и в тканевой жидкости — 60 мм рт.ст.; в притекающей артериальной крови оно составляет 40 мм рт.ст. Благодаря этому градиенту двуокись углерода движется из тканей в капилляры. В результате ее парциальное давление возрастает, достигая в венозной крови 46—48 мм рт.ст. Под влиянием высокого парциального давления часть двуокиси углерода физически растворяется в плазме крови.
Роль карбоангидразы. Большая же часть двуокиси углерода претерпевает химические превращения. Благодаря ферменту карбоангидразе она соединяется с водой, образуя угольную кислоту Н2СО3. Особенно активно эта реакция идет в эритроцитах, мембрана которых хорошо проницаема для двуокиси углерода.
Угольная кислота (Н2СО3) диссоциирует на ионы водорода Н + и гидрокарбоната (НСОз), которые проникают через мембрану в плазму.
Наряду с этим двуокись углерода соединяется с белковым компонентом гемоглобина, образуя карбоаминовую связь.
В целом 1 л венозной крови фиксирует около 2 ммоль двуокиси углерода. Из этого количества 10 % находится в виде карбоаминовой связи с гемоглобином, 35 % составляют ионы гидрокарбоната в эритроцитах, и оставшиеся 55 % представлены угольной кислотой в плазме.
Не нашли то, что искали? Воспользуйтесь поиском:
Лучшие изречения: Студент — человек, постоянно откладывающий неизбежность. 11142 —  | 7508 —
| 7508 —  или читать все.
или читать все.
источник
Аллостерическая регуляция сродства гемоглобина к кислороду 2,3-дфг
2,3-ДФГ снижает сродство гемоглобина к кислороду и, таким образом, повышает отдачу кислорода тканям. Если кровь израсходовала весь свой запас ДФГ, гемоглобин остается фактически насыщенным кислородом. При акклиматизации в условиях высокогорья содержание ДФГ в эритроцитах резко увеличивается. ДФГ является аллотерическим лигандом, так как связывается с гемоглобином в другом по сравнению с О2 участком. ДФГ встраивается в полость тетрамерной молекулы гемоглобина, полость образована остатками всех 4 протомеров.
В Т – форме (дезоксигенерированной) молекулы Hb имеются дополнительные связи, и поэтому размер центральной полости больше, чем в R – форме (дезоксигемоглобине). Поэтому ДФГ взаимодействует только с Т – формой стабилизируя её, путем образования связи между атомами кислорода ДФГ и тремя положительно заряженными группами в каждой из — цепей.
В легких при высоком парциальном давлении кислород взаимодействует с Hb, изменяется конформация белка, уменьшается центральная полость и ДФГ вытесняется из гемоглобина.
Вопрос 9. Реакции синтеза гема, регуляция синтеза.
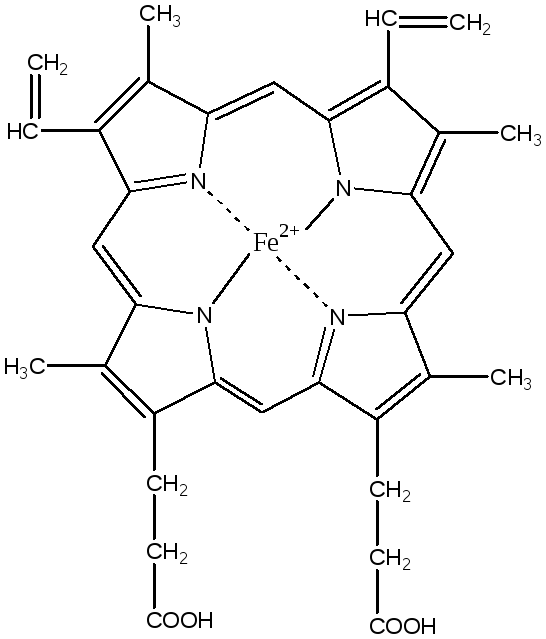
Гем является простетической группой гемоглобина, миоглобина, цитохромов, каталазы и пероксифазы.
Гем синтезируется во всех клетках, но наиболее активно синтез идет в печени и костном мозге, так как в этих тканях идет синтез гемоглобина и цитохромов. Гем в эритроцитах начинает синтезироваться уже на стадии полихроматофильных эритробластов. Клетки делится, постепенно накапливают гемоглобин, их цитоплазма становится оксифильной – это ацирофильные эритробласты. Синтез гема заканчивается только в зрелых эритроцитах (нет митохондрий).
Для синтеза гема необходимо два исходных реагента: глицин и сукцинил – КоА. Эти вещества необходимы для синтеза — аминолевулиновой кислоты ( — ALA). Реакция катализируется пиридоксальфосфатзависимым ферментам ALA – синтазой, которая локализуется в митохондриях.
Далее процесс перемещается в цитоплазму, так как следующий фермент синтеза гема – ALA – дегидротаза, локализован именно там. ALA – дегидротаза – Zn- содержащий фермент. Этот фермент катализирует реакцию конденсации двух молекул ALA с образованием пиррола – порфобилинагена (ПБГ).
Остальные стадии биосинтеза гема включают в себя соединение 4 молекул ПБГ в единую структуру, модификацию групп боковых цепей и образование хелатных комплексов с двух валентным атомом железа. Промежуточными тетрапехронами на стадиях между образованием ПБГ и формированием гема являются бесцветные (уро-) и копропорфириногемы. (содержащий метеновые мостики (-СН=)). Опишем подробнее стадии биосинтеза гема из ПБГ.
Уропорфириноген III превращается в копропорфириноген III путем декарбоксилирования всех ацетатных групп (А), вместо которых образуются метильные группы (М). Реакцию катализирует уропорфириноген – декарбоксилаза. Далее копропорфириноген III (КПГ) поступает в митохондрии, где превращается в протопорфириноген III (ППГ), а затем в протопорфирин III. Митоходриальный фермент копропорфириногеноксидаза катализирует декарбоксилирование и окисление двух пропионовых боковых цепей с образованием винильной группы во 2,4 положении (-СН=СН2). Так образуется ППГ. Затем он окисляется протопорфириногенооксидазой с образованием протопорфирина IX.
Завершающей стадией синтеза гема является включение в протопорфирин двух валентного железа. Эта реакция катализируется митохондриальным ферментом гем – синтезой или ферро – хелатазой.
Важным является и тот факт, что в процессе синтеза гема возрастает гидрофобность тетрапиррольных соединений в результате декарбоксилирования замещающих групп. Так, более полярный уропорфириноген экскретируется преимущественно с мочой, а более гидрофобные копропорфириноген и ППГ оказываются преимущественно в желчи и удаляются в составе фекалий.
источник
Сродство гемоглобина к кислороду. Эффект Бора
Эритроциты лишены митохондрий, поэтому в качестве энергетического материала они могут использовать только глюкозу. В эритроцитах катаболизм глюкозы обеспечивает сохранение структуры и функции гемоглобина, целостность мембран и образование энергии для работы ионных насосов. Глюкоза поступает в эритроциты путём облегчённой диффузии с помощью ГЛЮТ-2. Около 90% поступающей глюкозы используется в анаэробном гликолизе, а остальные 10% — в пентозофосфатном пути.
Конечный продукт анаэробного гликолиза лактат выходит в плазму крови и используется в других клетках, прежде всего гепатоцитах. АТФ, образующийся в анаэробном гликолизе, обеспечивает работу Nа + , К+-АТФ-азы и поддержание самого гликолиза, требующего затраты АТФ в гексокиназной и фосфофруктокиназной реакциях (см. раздел 7).
Важная особенность анаэробного гликолиза в эритроцитах по сравнению с другими клетками — присутствие в них фермента бисфосфоглицератмутазы. Бисфосфоглицератмутаза катализирует образование 2,3-бисфосфоглицерата из 1,3-бисфосфоглицерата (рис. 14-3). Образующийся только в эритроцитах 2,3-бисфосфоглицерат служит важным аллостерическим регулятором связывания кислорода гемоглобином (см. раздел 1).
Глюкоза в эритроцитах используется и в пентозофосфатном пути, окислительный этап которого обеспечивает образование кофермента NADPH, необходимого для восстановления глу-татиона
Метгемоглобинредуктаза представляет собой комплекс нескольких систем ферментов эритроцитов, катализирующих восстановление метгемоглобина, постоянно образующегося в небольшом (не более 1%) количестве в эритроците, в гемоглобин. Эти системы используют глюкозу и молочный сахар в виде источников Н+ и связаны с редуцированными формами НАДН2 и НАДФН2.
Большую роль в защите клетки от свободных радикалов играют ферментативные антиоксиданты. Эритроциты содержат высокоактивную супероксиддисмутазу, которая осуществляет дисмутацию двух O2 — с образованием перекиси водорода:
Образовавшаяся перекись водорода, являющаяся сильнейшим окислителем, частично нейтрализуется неферментативным путём при непосредственном участии аскорбата или других антиоксидантов (a-токоферол, глутатион восстановленный). Основное количество Н2О2 расщепляется в реакциях, катализируемых каталазой и глутатионпероксидазой:
Важную роль в антиоксидантной системе эритроцитов играют легкоокисляющиеся пептиды, содержащие аминокислоты с SH-группой: метионин, цистеин. Особое место занимает глутатион – трипептид, образованный цистеином, глутаматом, глицином. В организме он присутствует в окисленной и восстановленной форме (GSH). Основной антиоксидантный эффект глутатион оказывает, участвуя в работе ферментативных антиоксидантов. Глутатион является ингибитором активированных кислородных радикалов и стабилизатором мембран. Это связано с тем, что SH- содержащие соединения подвергаются окислению в первую очередь, что предохраняет от окисления другие функциональные группы.
Гемоглобин — сложный белок, хромопротеид, дыхательный пигмент крови человека, позвоночных и некоторых
беспозвоночных животных. Основная функция гемоглобина — перенос кислорода от органов дыхания к тканям.
Химически гемоглобин относится к группе хромопротеидов. Молекула гемоглобина состоит из белковой части —
глобина и простетичной группы небелковой природы — гема, в состав которого входит железо. В одной молекуле
гемоглобина содержится 4 простетических группы
Обратимое присоединение кислорода (оксигенация), позволяющее гемоглобину выполнять свою основную функцию переносчика, обеспечивается возможностью образовать прочные пятую и шестую координационные связи и перенести электрон на кислород не от железа (то есть окислить Fe2 +), а от имидазольного кольца проксимального гистидина. Это схематически изображено на рис. 1, б. Вместо молекулярного кислорода железо гема может присоединить окись углерода СО (угарный газ). Даже небольшие концентрации СО приводят к нарушению кислородпереносящей функции гемоглобина и отравлению угарным газом.
Выше было сказано, что одна молекула гемоглобина содержит четыре субъединицы и, следовательно четыре гема, каждый из которых может обратимо присоединить одну молекулу кислорода. Поэтому реакцию оксигенации можно разделить на четыре стадии:
Сродство гемоглобина к кислороду. Эффект Бора.
Молекула гемоглобина может находиться в двух формах — напряженной и расслабленной. Расслабленная форма гемоглобина имеет свойство насыщаться кислородом в 70 раз быстрее, чем напряженная. Изменение фракций напряженной и расслабленной формы в общем количестве гемоглобина в крови обусловливает S-образный вид кривой диссоциации оксигемоглобина, а следовательно, так называемое сродство гемоглобина к кислороду. Если вероятность перехода от напряженной формы гемоглобина к расслабленной больше, то возрастает сродство гемоглобина к кислороду, и наоборот. Вероятность образования указанных фракций гемоглобина изменяется в большую или меньшую сторону под влиянием нескольких факторов.
Основной фактор — это связывание кислорода с геминовой фуппой молекулы гемоглобина. При этом чем больше геминовых фупп гемоглобина связывают кислород в эритроцитах, тем более легким становится переход молекулы гемоглобина к расслабленной форме и тем выше их сродство к кислороду. Поэтому при низком Р02, что имеет место в метаболически активных тканях, сродство гемоглобина к кислороду ниже, а при высоком Р02 — выше. Как только гемоглобин захватывает кислород, повышается его сродство к кислороду и молекула гемоглобина становится насыщенной при связывании с четырьмя молекулами кислорода
Эффект рН и температуры на сродство гемоглобина к кислороду. Молекулы гемоглобина способны реагировать с ионами водорода, в результате этой реакции происходит снижение сродства гемоглобина к кислороду. При насыщении гемоглобина менее 100 % низкое рН понижает связывание кислорода с гемоглобином — кривая диссоциации оксигемоглобина смещается вправо по оси х. Это изменение свойства гемоглобина под влиянием ионов водорода называется эффектом Бора. Метаболически активные ткани продуцируют кислоты, такую как молочная, и С02. Если рН плазмы крови снижается от 7,4 в норме до 7,2, что имеет место при сокращении мыщц, то концентрация кислорода в ней будет возрастать вследствие эффекта Бора. Например, при постоянном рН 7,4 кровь отдавала бы порядка 45 % кислорода, т. е. насыщение гемоглобина кислородом снижалось до 55 %. Однако когда рН снижается до 7,2, кривая диссоциации смещается по оси х вправо. В результате насыщение гемоглобина кислородом падает до 40 %, т. е. кровь может отдавать в тканях до 60 % кислорода, что на 1/з больше, чем при постоянном рН.
Эффект 2,3-дифосфоглицерата (2,3-ДФГ) на сродство гемоглобина к кислороду. При некоторых физиологических состояниях, например при понижении Р02 в крови ниже нормы (гипоксия) в результате пребывания человека на большой высоте над уровнем моря, снабжение тканей кислородом становится недостаточным. При гипоксии может понижаться сродство гемоглобина к кислороду вследствие увеличения содержания в эритроцитах 2,3-ДФГ. В отличие от эффекта Бора, уменьшение сродства гемоглобина к кислороду под влиянием 2,3-ДФГ не является обратимым в капиллярах легких. Однако при движении крови через капилляры легких эффект 2,3-ДФГ на снижение образования оксигемоглобина в эритроцитах (плоская часть кривой диссоциации оксигемоглобина) выражен в меньшей степени, чем отдача кислорода под влиянием 2,3-ДФГ в тканях (наклонная часть кривой), что обусловливает нормальное кислородное снабжение тканей.
 Большую роль печень играет в обмене углеводов. Глюкоза, приносимая из кишечника по воротной вене, в печени превращается в гликоген. Благодаря высоким запасам гликогена печень служит основным углеводным депо организма. Гликогенная функция печени обеспечивается действием ряда ферментов и регулируется центральной нервной системой и 1 гормонами — адреналином, инсулином, глюкагоном. В случае повышенной потребности организма в сахаре, например, во время усиленной мышечной работы или при голодании гликоген под действием фермента фосфорилазы превращается в глюкозу и поступает в кровь. Таким образом, печень регулирует постоянство глюкозы в крови и нормальное обеспечение ею органов и тканей.
Большую роль печень играет в обмене углеводов. Глюкоза, приносимая из кишечника по воротной вене, в печени превращается в гликоген. Благодаря высоким запасам гликогена печень служит основным углеводным депо организма. Гликогенная функция печени обеспечивается действием ряда ферментов и регулируется центральной нервной системой и 1 гормонами — адреналином, инсулином, глюкагоном. В случае повышенной потребности организма в сахаре, например, во время усиленной мышечной работы или при голодании гликоген под действием фермента фосфорилазы превращается в глюкозу и поступает в кровь. Таким образом, печень регулирует постоянство глюкозы в крови и нормальное обеспечение ею органов и тканей.
Фруктоза включается в путь распада глюкозы на стадии триозофосфатов (см. рис. 9.32, а). С наследственной недостаточностью фруктозо- 1-фосфатальдола-зы (фермент 2 на схеме) связана врожденная непереносимость фруктозы. В этом случае при наличии в пище фруктозы в тканях накапливается фруктозо-1-фосфат (вследствие блока реакции 2), который ингибирует альдолазу фруктозо-1,6-бис-фосфата — фермент гликолиза и глюконеогенеза.
В результате нарушается и распад, и синтез глюкозы. Кроме того, фруктозо-1-фосфат ингибирует фосфорилазу гликогена. Эти причины приводят к появлению гипоглюкоземии после приема пищи, содержащей фруктозу. Болезнь обычно обнаруживается после перехода с грудного кормления на пищу, содержащую сахарозу, и проявляется приступами рвоты и судорог после еды. При устранении фруктозы из рациона дети развиваются нормально.Известно также наследственное нарушение обмена фруктоземия, вызываемое недостаточностью фруктокиназы. Поступающая в организм фруктоза не подвергается никаким изменениям, обнаруживается в крови и выводится с мочой. Каких-либо других симптомов при фруктоземии не наблюдали.
Включается в метаболизм путем превращения в глюкозо-1-фосфат.
На рис. 9.32, б»представлены пути превращений галактозы. Реакция, катализируемая УДФ-глюкозо-4-эпимеразой обратима, и используется как для синтеза УДФ-галактозы (и затем лактозы), так и для катаболизма галактозы. Лактоза синтезируется только в молочной железе женщин и только в период лактации.
В печени происходит важнейшее превращение жирных кислот, из которых синтезируются жиры, свойственные для данного вида животного. Под действием фермента липазы жиры расщепляются на жирные кислоты и глицерин. Дальнейшая судьба глицерина похожа на судьбу глюкозы. Его превращение начинается с участием АТФ и заканчивается распадом до молочной кислоты с последующим окислением до углекислого газа и воды. Иногда при необходимости печень может синтезировать гликоген из молочной кислоты.
основным местом образования плазменных пре-β-липопротеинов (липопротеины очень низкой плотности – ЛПОНП) и α-липопротеинов (липопротеины высокой плотности – ЛПВП) является печень.
Рассмотрим образование ЛПОНП. Согласно данным литературы, основной белок апопротеин В-100 (апо Б-100) липопротеинов синтезируется в рибосомах шероховатого эндоплазматического ретикулума гепатоцитов. В гладком эндоплазматическом ретикулуме, где синтезируются и липидные компоненты, происходит сборка ЛПОНП. Одним из основных стимулов образования ЛПОНП является повышение концентрации неэстерифици-рованных жирных кислот (НЭЖК). Последние либо поступают в печень с током крови, будучи связанными с альбумином, либо синтезируются непосредственно в печени. НЭЖК служат главным источником образования триглицеридов (ТГ). Информация о наличии НЭЖК и ТГ передается на мембранно-связанные рибосомы шероховатого эндоплазматического ретикулума, что в свою очередь является сигналом для синтеза белка (апо В-100). Синтезированный белок внедряется в мембрану шероховатого ретикулума, и после взаимодействия с фосфолипидным бислоем от мембраны отделяется участок, состоящий из фосфолипидов (ФЛ) и белка, который и является предшественником ЛП-частицы. Далее белокфосфо-липидный комплекс поступает в гладкий эндоплазматический ретикулум, где взаимодействует с ТГ и эстерифицированным холестерином (ЭХС), в результате чего после соответствующих структурных перестроек формируются насцентные, т.е. незавершенные, частицы (н-ЛПОНП). Последние поступают через тубулярную сеть аппарата Гольджи в секреторные везикулы и в их составе доставляются к поверхности клетки, после чего очень низкой плотности (ЛПОНП) в печеночной клетке.
Рассмотрим роль печени в обмене стероидов, в частности холестерина. Часть холестерина поступает в организм с пищей, но значительно большее количество его синтезируется в печени из ацетил-КоА. Биосинтез холестерина в печени подавляется экзогенным холестерином, т.е. получаемым с пищей.
Таким образом, биосинтез холестерина в печени регулируется по принципу отрицательной обратной связи. Чем больше холестерина поступает с пищей, тем меньше его синтезируется в печени, и наоборот. Принято считать, что действие экзогенного холестерина на биосинтез его в печени связано с торможением β-гидрокси-β-метилглутарил-КоА-редуктазной реакции:

Часть синтезированного в печени холестерина выделяется из организма вместе с желчью, другая часть превращается в желчные кислоты и используется в других органах для синтеза стероидных гормонов и иных соединений.
В печени холестерин может взаимодействовать с жирными кислотами (в виде ацил-КоА) с образованием эфиров холестерина. Синтезированные в печени эфиры холестерина поступают в кровь, в которой содержится также определенное количество свободного холестерина.
Метаболизм билирубина представлен на рисунке.
• Высвобождение гема. Более 80% гема образуется в результате разрушения эритроцитов и около 20% — миоглобина и цитохромов.
• Протопорфирин -> биливердин. Трансформация протопорфирина гема в биливердин происходит под влиянием микросомальных оксидаз гепатоцитов.
• Биливердин -> билирубин. Окисление биливердина (катализируемое цито-зольной биливердинредуктазой) с образованием непрямого билирубина. Непрямой билирубин, циркулирующий в крови, связан с альбуминами и поэтому не фильтруется в почках и отсутствует в моче.

Основные этапы метаболизма билирубина
• Диссоциация комплекса «неконъюгированный билирубин—альбумин» и транспорт билирубина в гепатоциты. В их цитозоле билирубин образует комплекс с белками и глутатион-8-трансферазами.
• Диглюкуронизация билирубина в гепатоцитах с образованием растворимого в воде конъюгированного билирубина. Прямой билирубин не связан с альбумином. В связи с этим он активно («прямо») взаимодействует с диазореактивом Эрлиха, выявляющим этот пигмент.
• Экскреция конъюгированного билирубина в желчевыводящие пути.
• Трансформация конъюгированного билирубина:
+ в уробилиноген (в верхнем отделе тонкой кишки), всасывающийся в тонкой кишке и попадающий по системе воротной вены в печень, где разрушается в гепатоцитах.
+ в стеркобилиноген (в основном в толстой кишке).
— Часть стеркобилиногена всасывается в нижнем отделе толстой кишки и с кровью геморроидальных вен попадает в общий кровоток. Стеркобилиноген хорошо растворим в воде, не связан с белками и поэтому фильтруется в почках в мочу (придавая ей в норме соломенно-жёлтый цвет).
— Другая часть стеркобилиногена выделяется с экскрементами, окрашивая их.
У большинства людей в течение жизни бывают нарушения пигментации. К счастью, в основном они доброкачественные, ограничены и обратимы. Например, наиболее часто нарушения пигментации встречаются при разрешении воспалительных дерматозов в виде гипер- или гипопигментации и существуют в течение нескольких месяцев.
2 основных типа нарушения пигментации.
Лейкодерма и меланодерма. При лейкодермеотмечаются более светлые участки по сравнению с нормальной кожей, при меланодерме— более темные. Кроме того, нарушения пигментации подразделяются с учетом механизма их образования — повреждения меланоцитов или изменения содержания пигмента в коже.
Желтуха (болезнь Госпела) (лат. icterus) — желтушное окрашивание кожи и видимых слизистых оболочек, обусловленное повышенным содержанием в крови и тканях билирубина.
Классификация желтух
В зависимости от вида нарушений метаболизма билирубина и причин гипербилирубинемии можно выделить три типа желтух: желтуху гемолитическую (надпеченочную), желтуху паренхиматозную (печёночную) и желтуху механическую (подпечёночную).
- Надпечёночные желтухи — возникают в связи с усилением процесса образования билирубина. При этом повышается его непрямая (неконъюгированная) фракция.
- Печёночные желтухи. Развитие печеночных желтух связано с нарушением потребления (захвата) билирубина гепатоцитами. При этом повышается непрямая (неконъюгированная) фракция билирубина.
- Подпечёночные желтухи — возникают при нарушении оттока желчи по внепеченочным желчным протокам (обтурационная желтуха).
Клиника желтух
Желтуха — симптомокомплекс, представляющий собой окрашивание в жёлтый цвет кожи, склер, слизистых оболочек. Интенсивность окрашивания может быть совершенно разной — от бледно-жёлтого цвета до шафраново-оранжевого. Умеренно выраженная желтуха без изменения цвета мочи характерна для неконъюгированной гипербилирубинемии (при гемолизе или синдроме Жильбера). Более выраженная желтуха или желтуха с изменением цвета мочи свидетельствует о гепатобилиарной патологии. Моча у пациентов с желтухами приобретает тёмный цвет вследствие гипербилирубинемии. Иногда изменение окраски мочи предшествует возникновению желтухи.
Лабораторные тесты
Ни один тест не может отдифференцировать каждый из вариантов желтухи, но есть общие комбинации печеночных тестов, которые помогают определить локализацию.
| Надпеченочная желтуха | Печеночная желтуха | Подпеченочная желтуха | |
| Общий билирубин | Норма / Увеличен | Увеличен | Увеличен |
| Неконьюгированный билирубин | Увеличен | Норма / Увеличен | Норма |
| Коньюгированный билирубин | Норма | Увеличен | Увеличен |
| Уробилиноген | Увеличен | Норма / Увеличен | Снижен / Отрицательный |
| Цвет мочи | Норма | Тёмный | Тёмный |
| Цвет стула | Норма | Обесцвеченный | Обесцвеченный |
| Щелочная фосфатаза | Норма | Норма | Увеличен |
| АЛТ и АСТ | Норма | Увеличен | Норма |
Детоксикационная функция печени состоит в обезвреживании в результате происходящих в печени процессов биосинтеза ядовитых для человеческого организма веществ. Иногда они становятся безвредными или даже нейтральными органическими соединениями, чаще всего – белковыми
Как осуществляется обезвреживающая функция печени
1. путем включения токсических веществ в синтез безвредных для организма соединений (например, аммиак — в мочевину или нуклеиновые кислоты),
2. окислительными процессами (дегидрирование этанола под действием алкогольдегидрогеназы);
3. восстановительными процессами (нитросоединения превращаются в аминосоединения);
4. путем гидролиза (лекарственные вещества);
5. путем конъюгации с различными обезвреживающими веществами:
глюкуроновой кислотой;
серной кислотой;
глицином;
таурином;
цистеином;
уксусной кислотой;
6. путем метилирования (производные пурина).
Микросомальные оксидазы — ферменты, локализованные в мембранах гладкого ЭР, функционирующие в комплексе с двумя внемитохондриальными ЦПЭ. Ферменты, катализирующие восстановление одного атома молекулы О2 с образованием воды и включение другого атома кислорода в окисляемое вещество, получили название микросомальных оксидаз со смешанной функцией или микросомальных монооксигеназ. Окисление с участием монооксигеназ обычно изучают, используя препараты микросом.
Реакции конъюгации составляют вторую фазу метаболизма жирорастворимых ксенобиотиков, которые в первой фазе гидроксилировались или получили иные нуклеофильные группы при помощи микросомных монооксигеназ. Химические же соединения, которые уже имеют в своем составе реакционноспособные группы (ОН, СООН, NH2, SH), сразу вступают в реакции конъюгации без предварительных превращений с участием оксидаз со смешанными функциями. У млекопитающих наиболее распространены следующие реакции конъюгации: глюкуронидная, сульфатная, с глутатионом, с глутамином, с аминокислотами, метилирование, ацетилирование, гликозидирование. При этом чаще всего и не только у млекопитающих встречается конъюгация с глюкуроновой кислотой.
источник